Arv, miljø og dysleksi - Metoder, hovedfunn og implikasjoner for praksis
Artikkelen tar for seg hvilke metoder som brukes for å studere forholdet mellom arv og miljø og dysleksi og avkoding, hovedfunn fra slike studier og eventuelle praktiske implikasjoner fra disse funnene.
Formålet med artikkelen er å redegjøre for hvilke metoder som brukes for å studere forholdet mellom arv og miljø i utviklingen av dysleksi og ordavkoding, samt hovedfunn og praktiske implikasjoner fra slike studier. For familierisiko og dysleksi presenteres en metaanalyse av studier av barn hvor en av foreldrene har dysleksi. Den viser at barn av dyslektikere har i gjennomsnitt 38–50 % risiko for selv å utvikle denne vansken. Når det gjelder tvillingstudier, presenteres en metaanalyse som viser at hovedtendensen er at leseferdigheter og dysleksi har en arvelig komponent på rundt 50 %, mens det øvrige kan tilskrives miljørelaterte faktorer. Når det gjelder molekylærgenetiske studier, har en nylig publisert metaanalyse (Poelmans, Buitelaar, Pauls & Franke, 2011) funnet 10–14 ulike gener som assosieres med dysleksi. Resultatene impliserer at man ved utredning av lesevansker bør gjøre en grundig kartlegging av slike vansker i nær familie. Selv om det er en betydelig genetisk komponent, viser også tidligere studier at intervensjoner kan ha en svært god effekt.
Nature, nurture and dyslexia: Methods, main findings and implications for practice
The topic of interest in this paper is the relationship between nature and nurture in the development of word reading skills. The aim here is to look closely into the methods that can be used to study this relationship and present the main findings and consequences for practitioners from such studies. As for studies that have examined children with a familial risk of dyslexia, a synthesis of the results show that those children with a parent with dyslexia have on average 44 % chances of developing dyslexia. When it comes to twin studies, a main finding from such studies is that about 50 % of reading skills and dyslexia can be accounted for by heritability, and the remaining can be explained by environmental factors. Finally, in a brief review of studies based on molecular genetics, the results from a recent published meta-analysis (Poelmans, Buitelaar, Pauls & Franke, 2011) show that findings which link dyslexia to specific genes have been replicated for 10–14 different gene locus.
Innledning
Å utvikle gode leseferdigheter er en viktig del av barns utvikling. Dessverre vet vi at mange barn i større eller mindre grad strever med å lære seg dette, og det fører til store konsekvenser for videre skolegang og deltagelse i samfunnslivet. Å forstå prosessene som ligger til grunn for å utvikle funksjonelle leseferdigheter er derfor av avgjørende betydning både for det enkelte barn, men også i et samfunnsperspektiv. Fokuset for denne artikkelen er komponenten i leseferdigheter som er relatert til ordavkoding. Avkodingsferdigheter innebærer å kjenne sammenhengen mellom lyd og bokstav, slik at dette kan settes sammen til hele ord. Dysleksi diagnostiseres som regel dersom barnets avkodingsferdigheter er betydelig svakere enn det man skulle forvente ut fra alder og/eller evnenivå (Diagnostic and Statistical Manual of Mental Disorders, American Psychiatric Association [DSM IV], 1994; International Statistical Classification of Diseases and Related Health Problems 10th Revision [ICD-10], WHO, 1993). Gode avkodingsferdigheter er en forutsetning for å forstå innholdet i tekst man leser, og det er derfor avgjørende for de fleste teoretiske skolefag.
Allerede tidlig i leseforskningens historie dannet man seg hypoteser om at dysleksi også var arvelig, siden dyslektiske vansker i noen familier kunne bli observert over flere generasjoner (Pennington & Smith, 1988). Debatten om hvordan arv og miljø påvirker barns utvikling er en av de eldste, men også en av de mest kontroversielle diskusjonene innenfor pedagogikk og psykologi (Rutter, 2002). Dessverre er det ofte slik at når spørsmålet om forholdet mellom arv og miljø kommer opp, blir det redusert til en svært forenklet konklusjon om at barns utvikling er et resultat av et samspill mellom arv og miljø, uten noen nærmere konkretisering. Denne upresisheten bunner nok delvis i at konklusjonene som har blitt trukket fra atferdsgenetiske studier som har forsøkt å klargjøre forholdet mellom arv og miljø har vært svært omstridte (se Rutter, 2006 kap.1). Et stort problem har vært at enkelte forskere innenfor atferdsgenetikk aksepterte finansiering fra organisasjoner med rasistiske formål, og at resultatene har blitt mistolket og misbrukt av grupper med slike interesser (Rutter, 2006 kap.1). I tillegg ble en sentral forsker som gjorde pionerarbeid innenfor atferdsgenetikk senere mistenkt for forskningsjuks (se Mackintosh, 1995).
Det har også vært et problem at kommentatorer og populærvitenskapelige forfattere med stor gjennomslagskraft har overdrevet genetikkens rolle og således bidratt til å polarisere debatten (Harris, 1998; Pinker 2002). Hvis man imidlertid ser bort fra problemene dette fagområdet tidligere var befengt med, er det tydelig at man de siste årene har gjennomført studier med forbedrede metoder som bidrar med svært verdifull kunnskap av samspillet mellom arv og miljø i barns utvikling (se for eksempel Boomsma, Busjahn & Peltonen, 2002; Colledge, Bishop, Koeppen-Schomerus et al., 2002; Olson, Keenan, Byrne et al. 2011; Neale & McArdle, 2000; Rutter, 2006 kap.1).
Formålet med denne artikkelen er å redegjøre for hvilke metoder som brukes for å studere forholdet mellom arv og miljø, hovedfunn fra slike studier og eventuelle praktiske implikasjoner fra disse funnene. Når det gjelder oppsummering av hovedfunn fra disse studiene, vil jeg basere dette på metaanalyser. En metaanalyse er et kvantitativt sammendrag av studier som har til hensikt å gi en oversikt over et forskningsområde (Borenstein, Hedges, Higgins & Rothstein, 2009). I denne artikkelen vil jeg først presentere en egen metaanalyse av studier som har sett på familierisiko og dysleksi. Deretter vil jeg presentere en egen metaanalyse av tvillingstudier, og til slutt vil jeg se kort på en nylig publisert metaanalyse av studier som har brukt såkalte molekylærgenetiske design for å undersøke arvelighet av dysleksi. Før jeg går nærmere inn på metoder og resultater fra studiene, vil jeg imidlertid kort (og forenklet) redegjøre for mekanismene som ligger til grunn for at arv kan påvirke leseferdigheter.
Mekanismer til grunn for sammenhengen mellom arv og leseferdigheter
Det er en stor mengde studier som støtter at vansker knyttet til språkets lydsystem (fonologi) er en viktig årsak til dysleksi (Bus & van Ijzendoorn, 1999; Hulme & Snowling, 2009; Wagner & Torgesen, 1987). Den rådende forståelsen er at barn med dysleksi har en fonologisk vanske som medfører en svekket evne til å danne seg representasjoner i hjernen for lydene i språket (fonemer) (Hulme & Snowling, 2009; Snowling 2000). Selv om en viktig årsak til dysleksi er problemer med å danne fonologiske representasjoner av språklyder, er det viktig å understreke at utviklingen av leseferdigheter er best forstått som en svært kompleks prosess der mange faktorer virker inn på ulike nivåer, både genetisk, nevrologisk og miljøbetinget (Hulme & Snowling, 2009; Hulme, Snowling, Caravolas, & Carroll, 2005). Dersom et barn utvikler dysleksi, er det derfor sjelden en enkelt årsak til dette, men det er et samspill mellom ulike årsaker. I likhet med de fleste andre utviklingsforstyrrelser utvikles derfor også lesevansker gjennom et samspill av ulike risiko er og beskyttelsesfaktorer som opererer på ulike nivåer.
En viktig forutsetning for å utvikle funksjonelle leseferdigheter er at det dannes kretser i hjernen som består av en mengde nerveceller som kommuniserer med hverandre (Hulme & Snowling, 2009). Dannelsen av kretsene i hjernen er delvis et resultat av genetisk påvirkning og delvis et resultat av samspillet med omgivelsene. Geners påvirkning på dette er et resultat av en kompleks prosess (for oversikter se Flint, Greenspan & Kendler, 2010; Plomin & Kovas, 2005; Plomin & Crabbe, 2000; Rutter, 2006 kap.7). Gener kan påvirke hjernens utvikling fordi DNA, som er bærer av den genetiske informasjonen, spesifiserer mRNA, som igjen spesifiserer produksjonen av proteiner (Rutter, 2006 kap.7). Disse proteinene kan påvirke strukturer som utvikles i hjernen og hvordan disse fungerer. Akkurat hvordan dette skjer og hvordan disse proteinene virker inn og danner grunnlaget for en bestemt atferd eller ferdighet, vet vi foreløpig lite om (Rutter, 2006 kap.7). Det som kompliserer bildet ytterligere, er at DNA i alle celler er hovedsakelig likt. Det som bestemmer om en celle blir en blodcelle eller en hjernecelle, er hvilke gener i cellen som aktiveres. I tillegg til at denne aktiveringen er influert av DNA-relaterte faktorer, kan miljøpåvirkning deaktivere eller aktivere gener som er involvert i å kontrollere hjernens utvikling og funksjon (Rutter, Moffitt, & Caspi, 2006; Rutter 2007). Dette betyr altså at selv om årsakskjeden starter med gener og DNA, har gener ingen direkte effekt på leseferdigheter. Det som skjer, er at genetisk informasjon gjennom en kjede av prosesser kan påvirke strukturer som dannes i hjernen og hjernens funksjon, og således påvirke leseferdigheter indirekte, men denne prosessen er også influert av miljøpåvirkninger (hvor miljø i tillegg til psykologiske prosesser også omfatter ernæring, eksponering for kjemiske stoffer, temperatur osv.). Det er i tillegg et komplisert samspill mellom flere ulike gener som påvirker utviklingen av leseferdigheter, og det er ikke slik at det finnes ett bestemt gen «for» leseferdigheter (Rutter, 2006 kap.8).
Metoder og hovedfunn fra studier av arv, miljø og dysleksi
Studier av arvelighet og dysleksi, og forholdet mellom arv, miljø og dysleksi kan deles i to hovedtyper studier; atferdsgenetiske studier og molekylærgenetiske studier (Rutter, 2006, se også Hensler, Schatschneider, Taylor & Wagner, 2010; Plomin & Walker, 2003). Atferdsgenetiske studier undersøker arv og forholdet mellom arv og miljø basert på en eller annen form av naturlig eksperiment, enten ved at man undersøker hvordan en bestemt vanske nedarves i familier, eller studerer tvillinger (Rutter, 2006 kap.3). Til forskjell fra atferdsgenetiske studier er molekylærgenetiske studier basert på identifikasjon av de genene som ligger til grunn for arvelighet som er observert i atferdgenetiske studier (Rutter, 2006 kap. 3). En forutsetning for å forstå forholdet mellom arv, miljø og dysleksi, er at man gjør en riktig tolking av resultatene som de enkelte studietypene kan frembringe. Før jeg går videre til å si noe om de konkrete resultatene fra studiene, vil jeg derfor gjennomgå hver av disse undersøkelsestypene og belyse hvilke konklusjoner som potensielt kan (og ikke kan) trekkes fra disse.
Familiestudier
Gjennomføring og tolkning av resultater.
I familiestudier har man undersøkt familier hvor en av foreldrene har dysleksi og observert barna deres fra førskolelader og inn i skolealder for å se om de utvikler lesevansker eller dysleksi (se for eksempel Scarborough, 1989). For å kunne trekke konklusjoner fra slike studier er man avhengig av at utvalget av barn med familiær risiko for dysleksi sammenlignes med hvor hyppig dysleksi forekommer i en kontrollgruppe uten familiær risiko. Det er sentralt å merke seg at denne type studier ikke kan si noe om forholdet mellom arv og miljø, siden studiene ikke er basert på tvillinger. Det innebærer at studiene kan gi informasjon om sannsynligheten for at et barn skal utvikle dysleksi hvis en eller begge foreldrene har det, men kan ikke skille mellom i hvilken grad arv og miljø hver for seg bidrar til at dysleksi utvikles (Rutter, 2006 kap. 3). Likevel er disse studiene på flere måter nyttige for å observere forholdet mellom arv og dysleksi. For det første kan studiene bidra til å undersøke en hypotese om hvorvidt en vanske har en økt forekomst i bestemte familier. Dersom man finner at dette er tilfellet, øker det sannsynligheten for at vansken har en arvelig komponent (Pennington, 2002; Rutter, 2006 kap. 3). Funnene fra familiestudier kan også være avgjørende for å identifisere den såkalte fenotypen til en vanske. Når det gjelder dysleksi, vil identifisering av fenotypen innebære og avdekke kjernekarakteristika ved denne vansken (Hulme & Snowling, 2009). Denne informasjonen kan blant annet brukes til å lage kartleggingsinstrumenter som kan identifisere barn som er i risiko før vansken utvikler seg (Rutter, 2006 kap. 3; Pennington, 2002). Familiestudier kan også gi nyttig informasjon om komorbiditet, altså sannsynligheten for at dysleksi inntreffer sammen med andre vansker. Dersom man finner at dette er tilfelle, kan det danne en hypotese om at vanskene, når de opptrer sammen, skyldes et felles nedarvet trekk (Pennington, 2002). Samlet sett kan altså familiestudier gi svært verdifull informasjon som både kan bidra til å belyse forholdet mellom arv og dysleksi, men også i forhold til utredning og tidlig diagnostisering av vansker.
Metode for metaanalyse.
For å få en oversikt over familiestudier som er gjennomført om dysleksi, ble det gjort et litteratursøk i databasene PsycInfo, ERIC, PsycAPA og databasene i ISI. Nøkkelordene «at-risk» og «dyslexia» og «famil* risk» og «dyslexia» ble brukt i søket. For å bli inkludert i analysen måtte studiene ha undersøkt en gruppe barn av foreldre hvor en eller begge har dysleksi. Barna må være målt på avkodingsferdigheter etter at leseopplæringen starter og ha gjennomgått en utredning hvor det blir vurdert om de oppfyller kriteriene for en dysleksidiagnose sammenlignet med en kontrollgruppe. Ut fra dette måtte studien ha beregnet en prevalensskåre, det vil altså si hvor mange prosent av barna med familiær risiko som oppfylte kriteriene for en dysleksidiagnose.
Litteratursøket genererte 138 ulike treff. Av disse oppfylte 14 studier inklusjonskriteriene (se tabell 1). I tillegg til informasjon om prevalens ble også følgende informasjon kodet: utvalgsstørrelse, undersøkelsestype, hvordan foreldrene var valgt ut og diagnostisert, hvordan barna var diagnostisert og barnas alder ved diagnostisering. Informasjon fra studiene ble kodet i dataprogrammet «comprehensive metaanalysis» (Borenstein, Hedges, Higgins & Rothstein, 2005).
Tabell 1. Studier som undersøker forekomst av dysleksi hos barn av foreldre med dysleksi
|
STUDIE |
% AV BARN AV DYSLETIKERE SOM SELV UTVIKLER DYSLEKSI |
UTVALGS- STØRRELSE (ANTALL BARN I GRUPPEN MED DYSLEKSI DIAGNOSE) |
ALDER VED DIAGNOSTISERING |
DIAGNOSTISKE KRITERIER |
REKRUTTERINGS-REGIME FOR FORELDRE MED DYSLEKSI |
|
Blomert & Willems, 2010 |
44 % |
53 |
1. klasse |
Fått dysleksidiagnose etter utredning ved et senter for dysleksi |
Familierisiko ble avgjort gjennom et spørreskjema til foreldre om lesevansker i familien. |
|
Boets, Vandermosten, Poelmans et al. 2011 |
44 % |
16 |
I 1. og 3. klasse |
Skåre under 10. persentil på lesetest I 1. og 2. klasse |
Uklart hvordan foreldrene er valgt ut. |
|
Boets, Wouters, van Wieringen et al. 2007 |
29 % |
9 |
Slutten av 1. klasse |
Samleskår under -1.3 standardavvik på ordlesing, nonordlesing og staving |
Barna var valgt ut fordi minst en av foreldrene hadde dysleksi. Uklart hvordan deres dysleksi er diagnostisert. |
|
de Bree, Wijnen & Gerrits, 2010 |
50 % |
19 |
8:6 |
Samleskår under 1 standardavvik på staving ordlesing og nonordlesing |
Foreldrene ble testet på lesetester og non-verbal IQ, og måtte skåre under 10. persentil på en nonordlesetest eller ordlesingstest, eller under 25. persentil på begge. |
|
Elbro & Petersen, 2004 |
55 % |
47 |
7. klasse |
Under 20. persentil på ordlesing, nonordlesing |
Foreldrene ble testet med standardiserte tester, og måtte skåre -1 standardavvik under gjennomsnittet. |
|
Elbro, Borstrøm & Peterson, 1998 |
37 % |
18 |
2. klasse |
Under -1 standardavvik på nord og ordlesingstester. |
Foreldrene ble intervjuet, og deres dysleksi ble bekreftet med tester. |
|
Ho, Leung & Cheung, 2011 |
47 % |
36 |
3. året i førskole (i Hong Kong) |
Barn under gjennomsnittet ble klassifisert som svake lesere. |
Foreldre testet, og de med normal IQ og under -1 standardavvik fra gjennomsnittet på minst to lesetester ble klassifisert som dyslektikere. |
|
Pennington & Lefly, 2001 |
34 % |
22 |
Før og etter 2. klasse |
Utredet på to tidspunkter og klassifisert som dyslektikere basert på et avvik mellom skårer på lese- og stavetester og IQ. |
Frivillige familier som var medlemmer av en organisasjon for personer med lesevansker, eller hadde et eldre barn med lesevansker. |
|
Scarborough, 1989 |
42 % |
16 |
2. klasse |
Samlet skåre på lesetest under -2 standardavvik i forhold til forventet ut fra nonverbal IQ. |
Frivillige familier som ble diagnostisert med lesevansker basert på tester. |
|
Smith, 2009 |
58 % |
10 |
8:9 |
Barn med en standardskåre på under 85 på tre av seks lesetester ble klassifisert som barn med lesevansker. |
Foreldre som skåret under -1 standardavvik på flere lesetester. |
|
Snowling, Gallagher & Frith, 2003 |
66 % |
37 |
8 år |
Skåre på lesetest under -1 standardavvik |
Foreldre som skåret under -1 standardavvik på lese- og fonologisk bevissthetstester. |
|
Torppa, Lyytinen, Erskine et al. 2010 |
35 % |
37 |
Slutten av 2. klasse |
Under 10. persentil på tre av fire lesetester. |
En eller begge foreldre diagnostisert med lesevansker på bakgrunn av et stort testbatteri med lese- og evnetester. |
|
van Bergen, de Jong, Plakas et al. (manuskript i trykk) |
30 % |
42 |
Slutten av 2. klasse |
Under 10. persentil på en ordlesingstest |
Foreldrene ble testet på lesetester og non-verbal IQ, og måtte skåre under 10 persentil på en nonordlesetest eller ordlesingstest, eller under 20. persentil på begge. |
|
van Bergen, de Jong, Regtvoort, et al. 2011 |
32 % |
22 |
10:1 |
Under 10. persentil på en ordlesingstest |
Foreldrene ble testet på lesetester og non-verbal IQ, og måtte skåre under 10 persentil på en nonordlesetest eller ordlesingstest, eller under 20. persentil på begge. |
I metaanalysen ble det estimert en gjennomsnittlig effektstørrelse for benevningshastighet på tvers av studier. Den enkelte studie vektes da i henhold til utvalgsstørrelse; det vil si at studier med store utvalg tillegges mer vekt enn studier med små utvalg. Hvorvidt den gjennomsnittlige effektstørrelsen var forskjellig fra 0, ble signifikanstestet (z-test). I beregning og signifikanstesting av gjennomsnittlige effektstørrelser ble såkalte random effekt-modeller brukt (Borenstein, Hedges, Higgins & Rothstein, 2009).
Random effekt-modeller er basert på en antagelse om at forskjeller i effektstørrelser mellom studier ikke bare skyldes tilfeldige feil, men også skyldes reell variasjon mellom studier når det gjelder for eksempel hvilke aldersgruppe de undersøker, hvilke tester de bruker osv. Slike variabler som man tenker seg kan være viktig for å forklare forskjeller mellom studier, kalles for moderatorer. Moderatorer kan påvirke gruppeforskjeller i den enkelte undersøkelse, og dermed bidra til å forklare hvorfor studiene viser ulike resultater. For å avgjøre hvor store forskjeller det var mellom resultatene fra studiene, ble en signifikanstest for homogenitet (Q-test) benyttet. Dersom denne er signifikant er forskjellen mellom studier større enn det som kan tilskrives tilfeldige ulikheter. Det betyr altså at det sannsynligvis er reelle forskjeller mellom studiene som kan forklare hvorfor de viser ulike resultater. For å analysere variabler som kunne tenkes å ha innvirkning på hvorfor ulike studier fikk ulike resultater, ble det brukt en signifikanstest (Q-test) for å undersøke om gjennomsnittlig effektstørrelse varierte for subnett av studier inndelt i kategorier (for eksempel etter kuttpunkt for dysleksi).
Resultater fra metaanalyse
Figur 1 viser gjennomsnittlig prevalens for dysleksi hos barn av dyslektikere. Gjennomsnittlig prosentandel av barn av dyslektikere som selv utvikler dysleksi, er 44 % (konfidensintervall 38–50, p < .05). Som vi ser av figur 1 er det klart at det er stor variasjon når det gjelder hvor mange prosent av barna som oppfyller kriteriene for en dysleksidiagnose. Tabell 1 viser at studiene er gjennomført på svært ulike måter, og det kan påvirke resultatene. En analyse av forskjeller mellom studier viser at den er liten, og ikke større enn man ville forvente i et tilfeldig utvalg (Q (14) = 17.95, p = .16, I² = 27.55). Hvis vi likevel ser nærmere på hva som kan forklare forskjeller mellom studier, ser vi at kuttpunktet som den enkelte studie setter for å diagnostisere lesevansker hos barna, har betydning for hvor mange prosent som får en dysleksidiagnose.
For syv studier som har et kuttpunkt for dysleksi på under 10.. prosentil på leseferdigheter, er gjennomsnittlig prevalens 35 %, mens for syv studier med et mer liberalt kuttpunkt (< 15. prosentil) er gjennomsnittlig prevalens 53 %, og denne forskjellen er signifikant (Q(1) = 11.36, p < .001). I tillegg ser vi også en forskjell på prevalens hvis foreldrenes dysleksi er bekreftet med tester (gjennomsnittlig prevalens 45 %), enn hvis dysleksivurderingen utelukkende er basert på selvrapportering (gjennomsnittlig prevalens 40 %), men denne forskjellen er ikke signifikant (p > 0.05). Samlet sett ser vi at barn hvor en av foreldrene har dysleksi, har en betydelig økt risiko for selv å utvikle dysleksi. Det eksakte prosenttallet er imidlertid avhengig av metodevariasjoner i studiene, men er i området 38–50 %.
Figur 1. Prosentandel barn av foreldre med dysleksi som utvikler dysleksi (gjennomsnittlig prevalens vist med ♦, prevalens estimat for hver enkelt studie vist med ■.
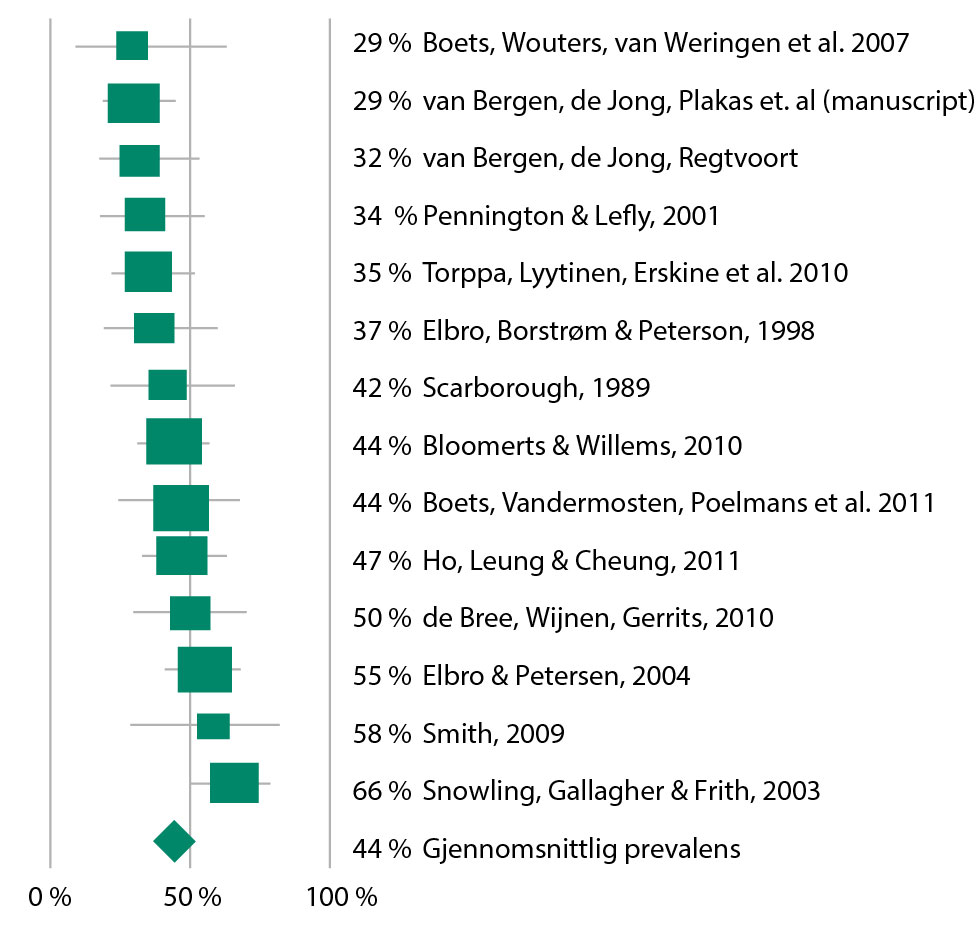
Det er viktig å merke seg at studier med dette designet ikke kan si noe om forholdet mellom arv og miljø, bare gi et estimat av en samlet risiko som er et resultat av både arv og miljøpåvirkning. I tillegg er det en viktig innvending mot denne type prevalensberegning at den er basert på at man setter et kuttpunkt for dysleksi, og klassifiserer barna i to grupper; de som utvikler dysleksi og de som ikke gjør det. Et slikt kuttpunkt vil være en kunstig grense, siden leseferdigheter er normalfordelt i populasjonen og barn med lesevansker varierer når det gjelder hvor alvorlige lesevanskene er (Hulme & Snowling, 2009). Hvis man ser nærmere på studiene av familierisiko og dysleksi, er det tydelig at barn av dyslektikere som ikke oppfyller kriteriene for en dysleksidiagnose, også har svekkete leseferdigheter sammenlignet med gjennomsnittet for kontrollgruppe uten arvelig risiko (se for eksempel Elbro & Petersen, 2004; Pennington & Lefly, 2001; Snowling, Gallagher & Frith, 2003). Studiene tyder altså på at det å ha en forelder med dysleksi også gir en økt risiko for å utvikle lesevansker av mindre alvorlig karakter, selv om man ikke nødvendigvis oppfyller kriteriene for en dysleksidiagnose. I fremtidige studier av familierisiko bør man derfor i større grad behandle leseferdigheter som en kontinuerlig variabel, fremfor å danne grupper basert på om barna med arvelig risiko utvikler dysleksi eller ikke (se Snowling, 2008).
Tvillingstudier
Gjennomføring og tolkning av resultater.
Tvillingstudier er basert på at man undersøker forskjeller mellom enog toeggede tvillingpar. Mens eneggede deler 100 % av arvematerialet, deler toeggede bare ca. 50 % av sine segregerende gener. Hvis de eneggede tvillingene er mer like på den ferdigheten eller egenskapen man studerer enn det toeggede tvillinger er, kan man trekke slutningen at dette skyldes at forskjellen mellom dem har oppstått som en følge av genetisk påvirkning (Rutter, 2006 kap.3). Tvillingstudier gjennomføres ofte ved at man tester tvillingene individuelt på mål som er interessante med hensyn til dysleksi (lesing, fonologiske ferdigheter med mer), og deretter beregner en såkalt intraklassekorrelasjon (for en oversikt over analysemetoder se Stromswold, 2001). Det er et mål for i hvilken grad personer innen en gruppe ligner på hverandre, og man beregner da en korrelasjon for de eneggede tvillingene og en for de toeggede tvillingene på hver av de ferdighetene man har målt. Hvis en bestemt ferdighet har en genetisk komponent, vil korrelasjonene mellom tvillingene være høyere for de eneggede enn for de toeggede. Forskjellen i korrelasjon mellom eneggede og toeggede tvillinger utgjør da et estimat for genetisk påvirkning. Det kan da beregnes en koeffisient for heritabilitet (2 ganger differansen i korrelasjonen mellom korrelasjonen for eneggede og korrelasjonen for toeggede tvillinger), som kan si noe om hvor mange prosent av variasjonen mellom barna i utvalget som kan tilskrives genetiske faktorer.
I tillegg til arvelighet finnes også variasjon som skyldes miljøpåvirkning. Variasjonen knyttet til miljøpåvirkning deles inn i to hovedtyper; delt miljøpåvirkning og unik miljøpåvirkning. Delt miljøpåvirkning inkluderer for eksempel den språklige påvirkningen tvillingene får fra foreldrene (gitt at foreldrene snakker på samme måte til begge), mens unik miljøpåvirkning er påvirkning som bare den ene i et tvillingpar opplever. Samlet sett gir altså en klassisk analyse av tvillingdata informasjon om tre komponenter, den arvelige komponent (A), en komponent som estimerer bidraget fra felles miljø (kalt C for common environment) og en komponent for den unike miljøpåvirkningen (E). En slik klassisk analyse av tvillingdata kalles derfor ofte for ACE-modellen (denne typen analyse annoteres som Falconer’s h² (Falconer& Mackay, 1996)).
Det bør også nevnes at ACE-modellen er den enkleste formen for analyse av tvillingdata. I takt med utviklingen av statistiske metoder generelt, har man også utviklet mer avanserte metoder for å analysere tvillingdata (se for eksempel Neale & McArdle, 2000). I slike analyser har man også mulighet til å ta høyde for at effekten av gener er avhengig av det miljøet de opererer i. I en analyse av gen/ miljøinteraksjon kan man estimere hvordan gener kan ha ulike utslag i ulike miljøer. I tillegg kan man også ta høyde for såkalt gen-miljøkorrelasjon, som innebærer at visse genvarianter opptrer oftere i bestemte miljøer. Hvis en gentype for eksempel gjør at en person liker å lese bøker, vil barn med denne genvarianten ha større sannsynlighet for å vokse opp i et hjem med flere bøker. Dette kan man undersøke ved å se på korrelasjonen mellom for eksempel antall bøker i hjemmet og genetisk variasjon.
En variant av tvillingstudien er at man undersøker tvillinger hvor en av dem har en lesevanske, og man ønsker å se på i hvilken grad vansken er arvelig. Da brukes en analyseform hvor man ser på hva som kan forklare forskjellen mellom grupper med og uten vansker fremfor hva som kan forklare variasjonen mellom personer (denne typen analyse annoteres ofte som DeFries & Fulker’s h²g (DeFries & Fulker, 1985)). Man finner da ut hvor mye av variasjonen mellom gruppene som kan tilskrives genetiske faktorer. Forskjellen mellom disse to metodene er altså at i den førstnevnte undersøker man hele variasjonen i populasjonens leseferdigheter, mens man i den sistnevnte deler inn populasjonen basert på de med lesevansker/dysleksi og normallesende kontrollbarn. I begge disse analysene vil man få ut estimat for arvelighet i tillegg til bidrag fra felles og unikt miljø.
For at slutningene man trekker fra en tvillingstudie skal være valide, ligger det en forutsetning til grunn om at forskjellen mellom eneggede og toeggede tvillingpar i sin helhet skyldes genetisk påvirkning, fordi miljøvariasjonen er lik for begge søsken innen hvert av de eneggede og de toeggede tvillingparene. Som Rutter (2006 kap. 3) påpeker, i utgangspunktet virker forutsetningen om lik miljøpåvirkning som umulig å oppfylle. Det vil jo være slik at eneggede tvillinger oftere går likt kledd, blir behandlet mer likt, og har likere interesser, og dermed får et likere miljø enn toeggede tvillinger. Siden dette imidlertid er en indirekte effekt av den genetiske påvirkningen, vil dermed ikke forutsetningen lik miljøvariasjon være brutt.
En svakhet ved tvillingdesignet som Rutter (2006, kap. 3) påpeker, er hvis det er en sterk genetisk effekt på en miljøvariabel som igjen har en sterk innvirkning på den egenskap eller atferd som studeres. Det er imidlertid viktig å huske på at dette bare er en svakhet dersom dette påvirker tvillingene i ulik grad, altså dersom den ene tvillingen for eksempel blir lest mye mindre for enn den andre, noe som kanskje er lite sannsynlig. Gitt et slikt scenario, vil imidlertid den genetiske påvirkningen bli overestimert (Rutter, 2006 kap.3).
En annen relevant svakhet er at tvillinger ikke er tilfeldig trukket fra en populasjon (Rutter 2006 kap.3). På grunn av det forutsetter generaliserbarheten fra studiene at tvillinger har omtrent den samme sannsynligheten som barn som ikke er tvillinger, til å utvikle en bestemt vanske og at tvillinger har et tilnærmet likt utviklingsforløp relatert til leseferdigheter som andre barn. Det er imidlertid ingenting som tyder på at tvillinger har høyere forekomst av språkvansker og dysleksi enn andre barn (Rutter, 2006 kap.3; Rutter & Redshaw, 1991; Thorpe, Rutter & Greenwood, 2003).
Samlet sett, i likhet med for alle studier, er det svakheter også med tvillingdesignet. Tvillingstudier regnes nå vanligvis, til tross for svakhetene og tidligere skepsis, for å være en valid metode for å undersøke påvirkning fra arv og miljø (Hensler, Schatschneider, Taylor & Wagner, 2010; Plomin & Walker, 2003; Rutter, 2006 kap.3).
Det er imidlertid viktig å huske på at estimatene fra tvillingstudiene gjelder utvalget som en helhet. Hvis man for eksempel finner et estimat for leseferdigheter som tilsier en arvelighet på 60 %, betyr det at forskjellene i leseferdigheter i det utvalget man har undersøkt, kan tilskrives genetiske faktorer. Man kan altså ikke overføre dette til et enkeltbarn og konkludere med at 60 % av vedkommendes lesevansker skyldes arv (Rutter, 2006 kap. 3). Et annet viktig moment er at dersom man finner at en egenskap har svært høy grad av arvelighet i en gruppe, betyr ikke dette at miljøet vil ha liten eller ingen innvirkning. Rutter (2006, kap. 3) eksemplifiserer dette med høyde, som har en arvelighet på ca. 90 %. Likevel har gjennomsnittshøyden økt vesentlig de siste 50 årene, sannsynligvis hovedsakelig på grunn av kostholdsendringer i populasjonen. Miljøet kan altså ha stor betydning selv om den prosentvise arveligheten man finner, er høy. Det er også viktig å merke seg at arveligheten i en ferdighet kan være veldig høy, selv om ferdigheten må læres og ikke er et konstant trekk. For eksempel ordforråd har en høy grad av arvelighet (se Stromswold, 2001) selv om hvert enkelt ord må læres.
Metode for metaanalyse.
For å få en oversikt over tvillingstudier som er gjennomført om dysleksi og leseferdigheter, ble det gjort et litteratursøk i databasene PsycInfo og ERIC. Nøkkelordene «twin*» og «dyslexia», «reading» og «decoding» ble brukt i søket. Litteratursøket gav 72 ulike treff. I tillegg ble det inkludert tvillingstudier rapportert i en metaanalyse av Stromswold (2001). For å bli inkludert i analysen, måtte studiene ha undersøkt enten tvillinger hvor minst en har dysleksi, eller leseferdigheter i grupper uten lesevansker. Studien måtte bestå av både eneggede og toeggede tvillinger. For å bli inkludert i analysen måtte studien rapportere et estimat for arvelighet (basert på en av de to metodene redegjort for ovenfor). For øvrig fulgte metaanalysen de metoder og prosedyrer som er beskrevet under forrige seksjon om familiestudier og dysleksi.
Resultater fra metaanalyse.
Resultatene fra tvillingstudiene er vist i Tabell 2. Når det gjelder resultatene fra studiene, er det verdt å merke seg at svært mange av undersøkelsene er basert på data fra den store Colorado-undersøkelsen. Dette er et såkalt kumulativt utvalg hvor tvillingpar blir inkludert i studien etter hvert som de blir født. Et problem ved dette er imidlertid at de ulike studiene i stor grad er basert på helt eller delvis overlappende utvalg, og dermed ikke uavhengig av hverandre. En forutsetning for metaanalyse er at man slår sammen data fra studier som er uavhengige observasjoner. En slik sammenslåing vil derfor for tvillingstudiene føre til et usikkert resultat, siden det i stor grad er overlapp mellom studiene. Hvis vi likevel gjør dette, er gjennomsnittlig arvelighetsestimat på 58 % [95 % Konfidensintervall 39.4 – 74.0] for de 9 studiene som har sett på hvor mange prosent av variasjonen mellom barna i utvalget som kan tilskrives genetiske faktorer (som har brukt Falconer’s h²). Ut fra Tabell 2 kan det også se ut som om studier fra Colorado-utvalget viser en høyere arvelighet enn de øvrige studiene. Det ble derfor gjort en ny beregning hvor bare den nyeste av Coloradostudiene var inkludert. Resultatet basert på 6 studier viste da en noe lavere arvelighet på 51 % [95 % KI = 27.28-75.74]. Når det gjelder de fire studiene som har undersøkt hvor mye av variasjonen mellom grupper med og uten lesevansker som tilskrives genetiske faktorer, er gjennomsnittlig arvelighetsestimat for dysleksi 55 % [95 % KI 44.2 –64.7]. Det understrekes imidlertid at disse gjennomsnittstallene bør tolkes svært forsiktig, siden de ikke er basert på uavhengige studier.
Som man kan se i Tabell 2, er det store forskjeller mellom studiene når det gjelder hvilket resultat de kommer frem til. Det er for få studier til å gjøre en statistisk analyse av faktorer som kan forklare forskjeller mellom studiene, men en viktig faktor som kan virke inn, er alvorlighetsgraden til lesevanskene: Blant annet Wadsworth, DeFries, Olson, og Willcutt (2007) har funnet at lesevansker av alvorlig karakter kan være mer arvelig enn leseferdigheter generelt. En annen viktig faktor som kan virke inn, er alderen til barna som blir undersøkt. For kognitive ferdigheter har man vist at arveligheten øker med økende alder, og er på sitt høyeste når ferdigheten er konsolidert i tidlig voksenalder (se for eksempel Haworth, Wright, Luciano et al., 2010). En mulig forklaring på dette er at etter hvert som barn blir eldre, velger og skaper de et miljø rundt seg som i større grad samsvarer med deres genetiske tilbøyeligheter enn det de har mulighet til i tidlig barnealder. Det fører til at den arvelige påvirkningen øker (Haworth, Wright, Luciano et al., 2010, se også Friend, et al., 2007).
I tillegg har det også vært vist at estimatene for arv av kognitive ferdigheter er lavere i barn som vokser opp i familier med lav sosioøkonomisk bakgrunn enn i de med høyere sosioøkonomisk bakgrunn (Turkheimer, Haley, Waldron, D’Onofrio & Gottesman, 2003). Det har blitt foreslått at dette skyldes at barn i familier fra lav sosioøkonomisk bakgrunn ikke fullt ut får utnyttet sitt genetiske potensial (Turkheimer, Haley, Waldron, D’Onofrio & Gottesman, 2003).
Samlet sett tyder altså denne analysen på at leseferdigheter har en genetisk komponent som forklarer rundt 50 % av variasjonen i populasjonen og mellom grupper med og uten lesevansker. Det innebærer altså at det også er en betydelig miljøpåvirkning som er avgjørende for utviklingen av leseferdigheter. Det er imidlertid behov for flere longitudinelle studier som grundig undersøker hvordan forholdet mellom arv og miljø utvikles over tid, og i tillegg også tar i betraktning viktige forhold som lesevanskenes alvorlighetsgrad og sosioøkonomisk bakgrunn.
Molekylærgenetiske studier
Gjennomføring og tolkning av resultat.
Molekylærgenetiske studier er en helt annen type studier enn de atferdsgenetiske, siden det her handler om å identifisere individuelle gener som kan bidra til å forklare arveligheten av en vanske eller et trekk (for oversikter se Rutter, 2006 kap. 8; Freimer & Sabatti, 2004). Slike studier er en forutsetning for å kunne endelig avklare arveligheten til en vanske og å lokalisere denne. Molekylærgenetiske studier kan deles i to hovedtyper, koplingsstudier og assosiasjonsstudier (Freimer & Sabatti, 2004). I koplingsstudiene fokuserer man som regel på søskenpar hvor begge har dysleksi. Deretter undersøker man om et utvalg søskenpar med dysleksi har like genetiske segmenter på sine kromosomer ved å kartlegge hele deres genetiske sammensetning.
Tabell 2. Tvillingstudier som undersøker forholdet mellom arv og miljø i utviklingen av lesevansker eller leseferdigheter.
|
STUDIE |
UTVALGSTYPE |
ESTIMAT FOR ARV |
UTVALGSALDER |
UTVALGSSTØRRELSE |
LOKALISERING |
|
Alarcon, & DeFries, 1997 |
Tvillinger med og uten lesevansker |
82 % på sumskåre leseferdigheter for tvillinger med lesevansker og 66 % for kontrolltvillinger uten lesevansker. (h²) |
|
276 eneggede og 210 toeggede (med samme kjønn) med lesevansker og 195 eneggede og 119 (med samme kjønn) uten lesevansker |
|
|
Boada, Willcutt, Tunick et al 2002 |
Tvillinger med gode leseferdigheter (ett standard-avvik over gjennomsnittet) |
55 % for sumskåre avkoding (h²g) |
11,53 |
215 eneggede og 135 toeggede med samme kjønn |
Fra Colorado utvalg |
|
Byrne, Samuelsson, Wadsworth et al. 2007 |
Uselekterte tvillinger |
82 % for samleskåre på Towre og 20 % for samleskår på CTOPP (fonologisk bevissthetstest) (h²) |
1. klasse |
167 eneggede og 152 toeggede |
197 fra Colorado utvalg |
|
Castles, Datta, Gayan & Olson, 1999 |
Tvillinger med dysleksi |
61 % for sumskåre avkoding (h²g) |
11,60 |
272 eneggede og 320 toeggede (hvorav 187 med samme kjønn ) |
Fra Colorado utvalg |
|
Casto, Pennington, Light & de Fries, 1996 |
Tvillinger hvor minst en hadde lesevansker |
35 % på en samleskår basert på leseferdigheter sett i forhold til evnenivå (h²g) |
fra 8–20 |
82 eneggede og 58 toeggede |
|
|
Hohonen & Stevenson, 1999 |
Tvillinger uten lesevansker (kryss-seksjonell studie med to aldersgrupper) |
60 % hos de yngre barna på en samleskåre for leseferdigheter, 59 % hos de eldre barna (h²) |
Yngre utvalg 5,10 år, elder utvalg 7,0 år. |
Yngre utvalg 32 eneggede og 28 toeggede, eldre utvalg 34 eneggede og 32 toeggede |
London |
|
Light, Defries, & Olson, 1998 |
Tvillinger med og uten lesevansker |
42 % for tvillinger uten lesevansker og 70 % for tvillinger med lesevansker på en sumskåre for leseferdigheter (h²) |
fra 8–20 |
132 eneggede og 91 toeggede (samme kjønn) uten vansker og 196 eneggede og 155 toeggede med vansker |
Colorado |
|
Olson, Keenan, Byrne et al 2011 |
Tvillinger uten lesevansker |
81 % for en samlevariabel for leseferdigheter (h²) |
2. klasse |
406 eneggede og 424 toeggede |
Colorado, Norge, Sverige, Australia |
|
Stevenson, Grahams, Fredman & McLoughlin, 1987 |
Tvillinger uten spesielle vansker |
18 % på lesetesten NEALE (h²) |
13,00 |
194 eneggede, 214 toeggede samme kjønn, 142 motsatt kjønn. |
London |
|
Thompson, Detterman & Plomin, 1991 |
Tvillinger uten spesielle vansker |
27 % på lesetesten Metropolitan Achievement test, reading (h²) |
6 til 13 |
146 eneggede og 132 toeggede |
Western reserve tvillingprosjekt. |
|
Wadsworth, Defries, Olson, Willcutt, 2007 |
Tvillinger med og uten lesevansker |
79 % for samleskåre lesing i gruppen med lesevansker (h²) og 36 % for samleskåre lesing i gruppen uten lesevansker (h²). |
10,40 |
106 eneggede og 172 toeggede, 124 tvillinger med lesevansker, 154 uten lesevansker |
Colorado |
|
Willcutt, Pennington et al 2007 |
Tvillinger med lesevansker |
66 % på en samleskåre basert på leseferdigheter sett i forhold til evnenivå (h²g) |
Informasjon mangler |
99 eneggede og 80 toeggede med lesevansker, |
Colorado |
Ved hjelp av statistisk analyse avgjør man deretter om likhetene man finner, er større enn de som oppstår ved tilfeldigheter. Ulempen er at for dysleksi er påvirkningen fra enkeltgener relativt svak (Rutter, 2006 kap. 8). Dette designet forutsetter derfor svært store utvalgsstørrelser for å oppdage effekter fra spesifikke gensammensetninger (Rutter, 2006 kap. 8; Freimer & Sabatti, 2004).
Den andre typen design er såkalte assosiasjonsstudier (for oversikt se Freimer & Sabatti, 2004). I disse studiene undersøker man om personer med dysleksi har større sannsynlighet enn kontrollbarn for å ha en bestemt genetisk sammensetning. Dette kan gjøres på to måter, enten ved at man starter med en hypotese om at et bestemt gen eller kombinasjon av gener er assosiert med dysleksi, og deretter prøver å finne en sammenheng, eller at man undersøker hele genomet og ser hvilke gener som viser sammenheng og skiller dyslektikerne fra kontrollpersonene (Freimer & Sabatti, 2004). Fordelen med dette er at man kan finne genetiske sammensetninger eller gener som har relativt små effekter på dysleksi. Ulempen er imidlertid at man ofte får falske positive funn, fordi det er vanskelig å avgjøre statistisk sannsynligheten for at en bestemt genkombinasjon har oppstått tilfeldig eller ikke (Rutter, 2006 kap. 8; Freimer
& Sabatti, 2004).
Resultater fra meta analysen til Poelmans, Buitelaar, Pauls og Franke (2011).
I en nylig publisert metaanalyse gjennomgår Poelmans, Buitelaar, Pauls og Franke (2011) koplingsstudier og assosiasjonsstudier som er gjort av dysleksi. En metaanalyse er spesielt interessant når det gjelder molekylærgenetiske studier, fordi det på grunn av metodiske problemer ofte har vist seg vanskelig å replikere funn fra tidligere studier (Rutter, 2006 kap. 8). Det innebærer at man altså har funnet utslag på gener i en studie, men at studier gjort i etterkant ikke har greid å bekrefte dette. Dermed sitter man igjen med et stort antall gener som er mulige kandidater som kan påvirke leseferdigheter og dysleksi. Dersom man analyserer dette ved hjelp av en metaanalyse, kan man sammenholde funn fra de ulike studiene, og dette vil gi en indikasjon på hvilke funn som er mest robuste.
Basert på en svært grundig metaanalyse finner Poelmans, Buitelaar, Pauls og Franke (2011) støtte for at dysleksi er relatert til 14 ulike gener, hvorav 10 kan plasseres i en teoretisk modell som beskriver et genetisk nettverk for hvordan disse genene påvirker dannelsen og migrasjonen av nerveceller. Det understrekes imidlertid at selv om man nå har gjort en rekke studier som har gitt kunnskap om sammenhengen mellom dysleksi og ulike gener, er det behov for nye studier for å verifisere hvordan disse genene virker sammen i et nettverk. Sannsynligvis finnes det flere slike forklaringsmodeller for hvordan gener relatert til dysleksi kan påvirke nerveceller.
Implikasjoner for pedagogisk og spesialpedagogisk praksis
I denne gjennomgangen av forskning på sammenhengen mellom arv, miljø og utviklingen av leseferdigheter og lesevansker, har vi sett at barn av foreldre med dysleksi har en risiko på mellom 38–50 % for å utvikle dysleksi. Når det gjelder tvillingstudier, er den genetiske komponenten for dysleksi og utvikling av leseferdigheter i overkant av 50 %. Dette tallet bør imidlertid tolkes med noe forsiktighet, siden det er få uavhengige studier. Det er imidlertid viktig å merke seg at det ser ut til å være stor grad av sammenheng når det gjelder funn fra de to studietypene. Vi har også sett at man gjennom en metaanalyse av molekylærgenetiske studier kan identifisere 14 aktuelle gener som er relatert til dysleksi som er replikert i ulike studier.
Samlet sett viser altså dette at man bør ta høyde for at utviklingen av dysleksi og lesevansker har en betydelig arvelig komponent. Det er imidlertid viktig å understreke at ett enkelt gen ikke fører til utviklingen av lesevansker, men det er assosiert med ulike gener og kombinasjoner av disse.
Hvilke praktiske implikasjoner har så denne kunnskapen? Et viktig moment er at for å kunne utøve god praksis må det ligge god forskning til grunn. Studier som samlet gir oss en forståelse av en vanske både på et genetisk, nevrologisk, kognitivt og atferdsmessig plan er derfor av avgjørende betydning. En viktig konsekvens av dette er betydningen av å kartlegge familiehistorien med hensyn til vansker når man utreder barn hvor det er mistanke om dysleksi og lesevansker. Siden studiene viser en betydelig genetisk påvirkning når det gjelder utviklingen av dysleksi, vil det å kartlegge familiehistorie kunne gi viktig informasjon om barnet muligens kan ha en genetisk risiko for lesevansker.
Det gir også viktig informasjon til foreldre som selv har dyslektiske vansker, om at man bør være oppmerksom på arveligheten av disse vanskene. Selv om prosenttallene fra studiene ikke kan trekkes ned på individnivå, vil det altså være en økt risiko for at barn arver slike vansker. Ved å følge med på barns språklige bevissthet og leseutvikling kan man sikre at barnet blir utredet og får hjelp i forhold til disse vanskene på et tidlig tidspunkt.
En annen viktig implikasjon av funnene er at selv om den genetiske påvirkningen er stor, kan intervensjoner ha god effekt for å avhjelpe vanskene. Det er også underbygget i en rekke treningsstudier som viser at man ved å arbeide med fonologisk bevissthet og bokstavkunnskap kan forbedre leseferdigheter betydelig (for oversikt se National Institute for Literacy 2008). Imidlertid, selv om det er interessant å vite prosentvis bidrag fra arv og miljø, har vel de praktiske konsekvensene av dette en noe begrenset betydning. Av større betydning for praksis er nok funnene fra familiestudiene. Disse har brakt oss nærmere en forståelse av fenotypen for dysleksi. På bakgrunn av denne kunnskapen kan vi derfor utvikle kartleggingsinstrumenter som kan fange opp barn med vansker på et tidlig tidspunkt samt lage målrettede intervensjoner som kan bidra til å avhjelpe vansken.
Litteraturhenvisninger
Alarcón, M., & Defries, J.C. (1997). Reading performance and general cognitive ability in twins with reading difficulties and control pairs. Personality and Individual Differences, 22, 793–803.
American Psychiatric Association (1994). Diagnostic and statistic manual of mental disorder (DSM IV). Washington DC: American Psychiatric Association.
Blomert, L. & Willems, G. (2010). Is there a causal link from a phonological awareness deficit to reading failure in children at familial risk for dyslexia? Dyslexia 16, 300–317.
Boada, R., Willcutt, E.G., Tunick, R.A., Chhabildas, N.A., Olson, R.K., Defries, J.C., & Pennington, B.F. (2002). A twin study of the etiology of high reading ability. Reading and Writing: An Interdisciplinary Journal, 15, 683–707.
Boets, B., Wouters, J., Van Wieringen, A., & Ghesquie`Re, P. (2007). Auditory processing, speech perception and phonological ability in preschool children at high-risk for dyslexia: A longitudinal study of the auditory temporal processing theory. Neuropsychologia, 45, 1608–1620.
Boomsma, D., Busjahn, A., & Peltonen, L. (2002). Classical twin studies and beyond. Nature Reviews Genetics, 3(11), 872–882.
Borenstein, M., Hedges, L. V., Higgins, J.P.T., & Rothstein, H. R. (2009). Introduction to meta-analysis. Chichester, UK: Wiley.
Borenstein, M., Hedges, L., Higgins, J., & Rothstein, H. (2005). Comprehensive Meta-Analysis Version 2 [software]. Engelwood, NJ: Biostat.
Bree, E.H. De, Wijnen, F.N.K. & Gerrits, P.A.M. (2010). Non-word repetition and literacy in Dutch children at-risk of dyslexia and children with SLI: Results of the follow-up study. Dyslexia, 16, 36–44.
Bus, A.G., & Van Ijzendoorn, M.H. (1999). Phonological awareness and early reading: A meta-analysis of experimental training studies. Journal of Educational Psychology, 3, 403 – 414.
Byrne, B., Samuelsson, S., Wadsworth, S., Hulslander, J., Corley, R., Defries, J. C., Quain, P., Willcutt, E. & Olson, R.K. (2007). Longitudinal twin study of early literacy development: Preschool through Grade 1. Reading and Writing: An Interdisciplinary Journal, 20, 77–102.
Casto, S.D., Pennington, B.F., Light, J.G., & Defries, J.C. (1996). Differential genetic etiology of reading disability as a function of mathematics performance. Reading and Writing: An Interdisciplinary Journal, 8, 295–306.
Castles, A., Datta, H., Gaya´N, J. & Olson, R.K. (1999). Varieties of developmental reading disorder: Genetic and environmental influences. Journal of Experimental Child Psychology, 72(2), 73–94.
Colledge, E., Bishop, D.V.M., Dale, P., Koeppen-Schomerus, G., Price, T. S., Happe, F. Et Al. (2002). The structure of language abilities at 4 Years: A twin study. Developmental Psychology, 38, 1–9.
Defries, J. & Fulker D. (1985). Multiple regression analysis of twin data. Behavior Genetics 15(5), 467–73.
Elbro, C., & Petersen, D. K. (2004). Long-term effects of phoneme awareness and letter name training. An intervention study with children at risk of dyslexia. Journal of Educational Psychology, 96(4), 660–670.
Elbro, C., Borstrøm, I., & Petersen, D. K. (1998). Predicting dyslexia from kindergarten: The importance of distinctness of phonological representations of lexical items. Reading Research Quarterly, 33, 36–60.
Falconer, D.S. & Mackay, T.F.C. (1995). Introduction to Quantitative Genetics (4th Edition). Addison Wesley Longman, New York.
Flint J., Greenspan R.J., Kendler K.S. (2010). How Genes Influence Behavior. Oxford: Oxford University Press.
Freimer, N. And C. Sabatti (2004). Pedigree, sib-pair, and association studies of common diseases; genetic mapping and epidemiology. Nature Genetics 36: 1045–1051.
Friend, A., Defries, J.C., Wadsworth, S.J. & Olson, R.K. (2007). Genetic and environmental influences on word recognition and spelling deficits as a function of age. Behavior Genetics, 37, 477–486.
Harris, J. (1998). The nurture assumption: Why children turn out the way they do. New York: Free Press
HAWORTH, C.M.A., WRIGHT, M.J., LUCIANO, M., MARTIN, N.G., DE GEUS, E.J.C., VAN BEIJSTERVELDT, C.E.M., BARTELS, M., POSTHUMA, D., BOOMSMA, D.I., DAVIS, O.S.P., KOVAS, Y., CORLEY, R.P., DEFRIES, J.C., HEWITT, J.K., OLSON, R.K., RHEA, S.A., WADS WORTH, S.J., IACONO, W.G., MCGUE, M., THOMPSON, L.A., HART, S.A., PETRILL, S.A., LUBINSKI, D. & PLOMIN, R. (2010). The heritability of general cognitive ability increases linearly from childhood to young adulthood. Molecular Psychiatry, 15, 1112–1120. doi: 10.1038/ mp.2009.55
Hensler, B.S., Schatschneider, C., Taylor, J., & Wagner, R.K. (2010). Behavioral genetic approach to the study of dyslexia. Journal of developmental behavioral pediatrics’, 31(7), 525–32.
Ho, K.S.H., Leung, M.T. & Cheung, H. (2011). Early difficulties of Chinese preschoolers at familial risk for dyslexia: deficits in oral language, phonological processing skills, and print-related skills. Dyslexia, DOI: 10.1002/dys.429.
Hohnen, B., And Stevenson, J. (1999). The Structure of Genetic Influences on General Cognitive, Language, Phonological, and Reading Abilities. Developmental Psychology 35, 590–603.
Hulme, C., Snowling, M., Caravolas, M., & Carroll, J. (2005). Phonological skills are (probably) one cause of success in learning to read: A comment on Castles and Coltheart. Scientific Studies of Reading. 9 (4) 351–365.
Hulme, C. & Snowling, M. (2009). Developmental Cognitive Disorders. Oxford: Blackwell/Wiley.
Light, J.G., Defries, J.C., & Olson, R.K. (1998). Multivariate behavioral genetic analysis of achievement and cognitive measures in readingdisabled and control twin pairs. Human Biology, 70, 215–237.
Mackintosh, N.J. (1995). Cyril Burt: Fraud or Framed? Oxford: Oxford University Press.
National Institute For Literacy (2008). Developing early literacy: A scientific analysis of early literacy development and implications for intervention. NIFL gov: http://www.nifl.gov/publications/pdf/NEL- PReport09.pdf
Neale, M.C., Mcardle, J.J. (2000). Structured latent growth curves for twin data. Twin Research, 3, 165–77. 147.
Olson, R. K., Keenan, J. M., Byrne, B., Samuelsson, S., Coventry, W. L., Corley, R.,Wadsworth, S. J., Willcutt, E. G., Defries, J. C., Pennington, B. F., & Hulslander, J. (2011). Genetic and environmental influences on vocabulary and reading development from pre kindergarten through grade 4. Scientific Studies of Reading, 15, 26–46.
Pennington, B. (2002). The development of psychopathology: nature and nurture. London: Taylor & Francis Books.
Pennington, B. F., & Lefly, D. L. (2001). Early reading development in children at family risk for dyslexia. Child Development, 72, 816–833.
Pinker, S. (2002). The blank slate: the modern denial of human nature. London: Penguin.
Plomin, R. And Walker, S. O. (2003), Genetics and educational psychology. British Journal of Educational Psychology, 73: 3–14.
Plomin, R. & Kovas, Y. (2005). Generalist genes and learning disabilities. Psychological Bulletin, 131 (4), 592–617.
Plomin, R. & Crabbe, J. (2000). DNA. Psychological Bulletin, 126, 806–828.
Poelmans, G., Buitelaar, J.K., Pauls, D.L., & Franke, B. (2011). A theoretical molecular network for dyslexia: integrating available genetic findings. Molecular Psychiatry 16 (4), 365–82.
Rutter, M., Moffitt, T.E. & Caspi, A. (2006). Gene-environment interplay and psychopathology: multiple varieties but real effects. Journal of. Child Psychology and Psychiatry, 47, 226–261.
Rutter M., & Redshaw, J. (1991): Annotation: growing up as a twin: twin-singleton differences in psychological development. Journal of. Child Psychology and Psychiatry 32, 885–895.
Rutter, M. (2002). Nature, nurture, and development: From evangelism through science towards policy and practice. Child Development, 73, 1–21.
Rutter, M. (2006). Genes and behavior: Nature– nurture interplay explained. Oxford: Blackwell Publishing.
Rutter, M. (2007). Gene-environment interdependence. Developmental Science, 7(10), 12–18.
Scarborough, H. S. (1989). Prediction of reading disability from familial and individual differences. Journal of Educational Psychology, 81, 101–108.
Smith, S.(2009). Auditory processing, speech perception and phonological ability in preschool children at high-risk for dyslexia. Reading and Writing 22, 1, 25–40.
Snowling, M. J. (2000). Dyslexia. Oxford, UK: Blackwell Publishers.
Snowling, M. J. (2008). Specific disorders and broader phenotypes: The case of dyslexia. Quarterly Journal of Experimental Psychology, 61, 142–156.
Snowling, M.J. Gallagher, A. & Frith, U. (2003). Family risk of dyslexia is continuous: individual differences in the precursors of reading skill. Child Development, 74, 358–373.
Stromswold, K. (2001). The heritability of language: A review and meta-analysis of twin, adoption and linkage studies. Language, 77(4), 647–723.
Stevenson, J, Graham, P, Fredman, G, & Mcloughlin, V. (1987), A twin study of genetic influences on reading and spelling ability and disability. Joumal of Child Psychology and Psychiatry, 28, 229–247.
Thompson, L. A., Detterman, D. K., & Plomin, R. (1991). Associations between cognitive abilities and scholastic achievement: Genetic overlap but environmental differences. Psychological Science, 2, 158 – 165.
Thorpe, K., Rutter, M., & Greenwood, K. (2003). Twins as a natural experiment to study the causes of mild language delay II: Family interaction risk factors. Journal of Child Psychology and Psychiatry, 44, 342–355.
Torppa, M., Lyytinen, P., Erskine, J., Eklund K. & Lyytinen H. (2010). Language development, literacy skills, and predicitive connections to reading in Finnish children with and without familial risk for dyslexia. Journal of Learning Disabilities, 43(4), 308–21.
Turkheimer, E., Haley, A., Waldron, M., D’onofrio, B.M., Gottesman, I.I. (2003). Socioeconomic status modifies heritability of IQ in young children. Psychological Science, 14, 623–628.
Van Bergen, E., De Jong, P.F., Plakas, A., Maassen, B. & Van Der Leij, A. (In press). Child and parental literacy levels within families with a history of dyslexia. Journal of Child Psychology and Psychiatry. In press van Bergen, E., de Jong, P.F., Regtvoort, A., Oort, F., S. van Otterloo, & van der Leij, A. (2011). Dutch children at family risk of dyslexia: Precursors, reading development and parental effects. Dyslexia, 17(1), 2–18.
Wadsworth, S.J., Defries, J.C., Olson, R.K., & Willcutt, E.G. (2007). Colorado Longitudinal Twin Study of Reading Disability. Annals of Dyslexia, 57, 137–160.
Wagner, R. & Torgesen, J. (1987). The nature of phonological processing and its causal role in the acquisition of reading skills. Psychological Bulletin, 101, 192 – 212.
Willcutt, E. G., Pennington, B. F., Olson, R. K., & Defries, J. C. (2007). Understanding comorbidity: A twin study of reading disability and attention-deficit/hyperactivity disorder. American Journal of Medical Genetics (Neuropsychiatric Genetics), 8, 709 – 714.
World Health Organization. (1993). The ICD-10 classification of mental and behavioral disorders. Diagnostic criteria for research. Geneva: World Health Organization.